Marco Arrigoni, Nigel Gallaher, Darryl McCoy, Volker Pfeufer and Matthais Schulze, Coherent Inc.
Laser development for the microscopy market continues to be driven by key trends in applications, which currently include superresolution techniques, multiphoton applications in optogenetics and other areas of neuroscience, and even a shift in multiphoton imaging toward preclinical and clinical usage.
In spite of its long history, optical microscopy – particularly laser-based microscopy – is a very dynamic field. New techniques continue to be developed, while existing techniques are being applied to new applications. For biologists, drug developers, clinical lab professionals and other scientists to fully exploit these new techniques and applications, parallel developments in laser technology are often required. This article will take a broad overview of three important trends in laser-based microscopy and examine how laser manufacturers are responding with products optimized to match the needs of these applications.
Superresolution microscopy – optical switching of fluorescent labels
To better understand the details of processes such as signaling and control of gene expression, biologists need to correlate molecular-level events with macroscopic structures and dynamics. This has fueled rapid growth in superresolution microscopy techniques, often referred to as nanoscopy, that go beyond the classical spatial resolution limit set by diffraction. This limit is about half the wavelength of light (i.e., for visible light, approximately 200 to 250 nm) in the X-Y plane and, in the case of confocal microscopy, about 500 nm in the Z direction. Most superresolution techniques use CW lasers, often with fast internal or external (on/off) modulation.
The Nobel Prize committee recognized the importance of superresolution microscopy by awarding the 2014 Nobel Prize in chemistry jointly to Dr. Eric Betzig, Dr. Stefan Hell and Dr. William Moerner for their pioneering work in developing superresolution microscopy techniques.
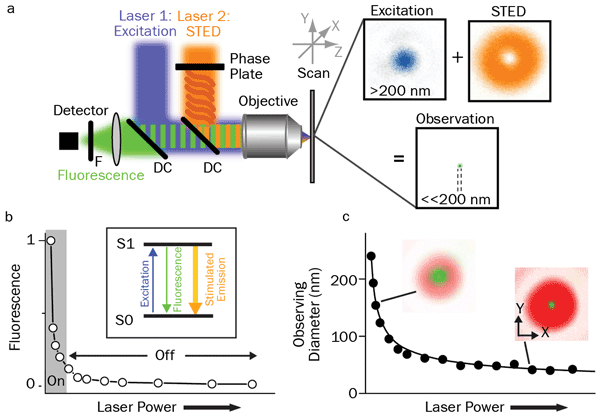
Figure 1. Principles of STED: (a) Schematic drawing of the setup of a STED nanoscope with phase plate, objective lens dichroic mirror (DC), fluorescence filter (F), detector, scanning device, and excitation and STED lasers with their focal intensity distribution (right) and a representative, subdiffraction-sized observation area. (b) STED nanoscopy is based on inhibiting fluorescence emission by de-exciting the excited S1 ‘On’ state to the S0 ‘Off’ ground state via stimulated emission. Increasing the power of the STED laser drives the inhibition into saturation. (c) Combined with an intensity distribution that features at least one intensity zero, this On/Off fluorescence inhibition realizes subdiffraction-sized observation volumes: The volume in which fluorescence emission is still allowed (green, insets) decreases with increasing STED laser power. Courtesy of Christian Eggeling; see acknowledgments.
All optical superresolution or nanoscopy techniques are based on the principle of optically and reversibly preparing states of a fluorescence label that differ in their emission characteristics (e.g., a bright “on” and a dark “off” state). Based on their different mechanisms for this on/off photoswitching, optical nanoscopy methods can be loosely divided into two groups: those that directly improve microscope effective spatial resolution by deterministic photoswitching in space and time, and those that achieve the higher resolution by (random) stochastically switching single-molecule fluorescence on and off in space.
Examples of the first group include stimulated emission depletion (STED) microscopy, ground-state depletion microscopy, reversible saturable optical fluorescence transition (RESOLFT) microscopy or parallelized RESOLFT microscopy, often denoted nonlinear or saturated structured illumination microscopy (SIM). Examples of the latter include direct stochastic optical reconstruction microscopy (d-STORM) and fluorescence photoactivation localization microscopy (f-PALM).
Figure 1 explains how STED works. A donut-shaped laser beam is coincident with a TEM00 (Gaussian profile) beam of a different wavelength. The donut beam depletes the population of excited fluorophores via stimulated emission. This beam shape serves to constrict the volume of fluorophores that the TEM00 beam excites. The donut-shaped depletion beam has a smooth spatial profile so that increasing the power in the STED beam increases the “bleached” area and thus confines fluorescence excitation by the TEM00 beam to a smaller area. This is equivalent to having a “knob” that controls the spatial resolution. Optimized on/off time-gating of the two lasers enables signal discrimination and high signal-to-noise images. Figure 2 is an example showing the effectiveness of STED.

Figure 2. The effectiveness of STED is clear in this pair of images of fluorescently tagged microtubuli in mammalian cells using a 577-nm Coherent Genesis laser to drive the STED effect: conventional confocal (left) and gated STED image (right). Image courtesy of Dr. Giuseppe Vicidomini and Dr. Alberto Diaspro, Istituto Italiano di Tecnologia (IIT), Genoa, Italy (G. Vicidomini, I.C. Hernández, M. d’Amora, F. Cella Zanacchi, P. Bianchini, A. Diaspro (2013). Gated CW-STED microscopy: A versatile tool for biological nanometer scale investigation. Methods, doi: 10.1016/j.ymeth.2013.06.029).
Stochastic-based techniques like f-PALM and d-STORM also use two lasers (Figure 3). Here, one laser drives most of the fluorophores between the “on” and “off” states, so that only a very small, random subset of well-separated molecules is in the “on” state and available to be excited by a second laser. The fluorescence spread from each of these camera-imaged point sources is then analyzed to find the centroid (i.e., the point source or spatial localization, based on the point spread function of the microscope). Repetitive on/off switching brings up different random subsets in subsequent camera images for single-molecule localization. After many cycles, the microscope computer assembles all the idealized point sources into a subdiffraction image, since the localization of a single molecule can be performed with high, subdiffraction spatial precision.
Trends in CW lasers to support superresolution techniques
Laser manufacturers are supporting the rapid growth in superresolution microscopy with new products that provide smoothly adjustable and higher power, new wavelengths, and simplified fiber coupling and combining.
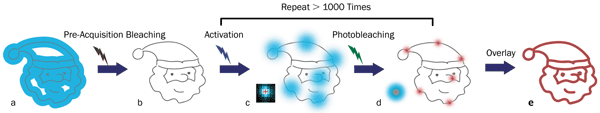
Figure 3. Principles of STORM microscopy: (a) A wide-field fluorescence image (blue area) is observed from overlapped fluorophores in a small target structure (gray lines); (b) no or little fluorescence is left after pre-acquisition bleaching with an appropriate laser; (c) individual fluorophores are sparsely activated by an activation laser and then emit fluorescence (blue spots) after applying an excitation laser (usually the same one for pre-acquisition bleaching). (d) The excitation laser is again used to bleach the fluorophores. The center positions (red crosses) of the fluorophores are found using an appropriate molecule-localization algorithm. The uncertainty of the center positions (red spots) is significantly smaller than the diameter of the single-molecule fluorescence from the same fluorophores (the large blue spot in the left corner). (e) Repeating the activation, excitation, localization and photobleaching process more than 1000 times allows reconstruction (via overlaying the center positions of the fluorophores) of a superresolution image of the target structure (red lines). Image courtesy of Dr. Zhen-li Huang at Wuhan National Laboratory for Optoelectronics, Huazhong University of Science and Technology.
Higher laser power is necessary because researchers want output in the 1- to 3-W range for STED and, typically, a few hundred milliwatts for PALM and STORM. In the case of SIM, microscope manufacturers are looking to move up to 300-mW lasers to decrease image acquisition times, whereas early users of this method worked with as little as 100 mW. Both deterministic and stochastic switching of fluorescence techniques need adjustable power output in the “bleaching” laser for optimization.
Lasers based on optically pumped semiconductor laser (OPSL) technology are proving ideal to meet these higher-/adjustable-power needs. That’s because OPSL technology is readily scalable, simply by increasing the pump diode power. And OPSL output can be smoothly adjusted from a few percent to maximum power with no effect on beam quality or beam pointing, since OPSL technology does not suffer from thermal lensing, which prevents wide power adjustment in diode-pumped and other solid-state lasers.
Another key trend is the growing demand for “new” – and particularly longer – laser wavelengths to optimally excite many new fluorophores. These include the mFruit series of fluorescent proteins and a host of new green, yellow and orange excited dyes originally developed for multichannel flow cytometry applications. As with wide-field microscopy, new nanoscopy methods often use combinations of multiple fluorophores to permit simultaneous mapping or probing for more than one type of structure or biochemical.
Fortunately, wavelength scalability is another key advantage of OPSL technology; lasers can be fabricated for any arbitrary output wavelength across much of the visible and near-UV spectrum, merely by tweaking the design of the gain semiconductor chip. For example, new lasers at 588 and 594 nm were recently introduced for exciting orange fluorophores. Other models at 550 nm have also been developed, as this wavelength can be used to excite green dyes and several new yellow fluorophores simultaneously with a single laser. Yet another new OPSL wavelength is the legacy (krypton ion) wavelength of 568 nm.
Fiber coupling/fiber combining
Many end users and microscopy OEMs – and not just in superresolution – are increasingly using fiber delivery with CW lasers. Maintaining high beam quality necessitates using polarization-preserving, single-mode fiber with a core diameter around 3.5 µm. At first, users would couple the laser into the single-mode fiber using mounts with up to six degrees of freedom that had to be individually optimized and then locked. This typically required many hours (per laser) from a skilled technician, and the fiber often lasted only hundreds of hours because of degradation of the small output facet. This problem was eliminated with the advent of bioinstrumentation lasers with permanent fiber-coupled outputs. And in the particular case of Coherent’s OBIS FP lasers, a proprietary fiber avoids rapid fiber degradation while preserving single-mode output.
The past few years have seen a similar paradigm shift in combining multiple lasers in response to the needs of applications to use multiple wavelengths simultaneously (see, for example, “Laser Combiner Enables Scanning Fluorescence Endoscopy,” November 2014 BioPhotonics). This formerly required multiple optics (beamsplitters, wave plates, etc.) for each laser wavelength and demanded hours of painstaking adjustment. But a new plug-and-play combiner, Coherent’s OBIS Galaxy, now simplifies this process. This passive device can integrate up to eight different laser wavelengths using a patent-pending combination of a minimum number of refractive optical components, together with a novel fiber having constant NA over the entire 405- to 640-nm range. This reduces the task to simple plug-and-play with snap-in (FC/UFC) fiber connectors.
Neuroscience dominates growth in multiphoton imaging
In multiphoton excitation (MPE) microscopy using ultrafast lasers, the single biggest driver in techniques and laser development is neuroscience. Billions of dollars in public and private funding is enabling dramatic advances in understanding how the brain processes information, from the synaptic to the network level. Moreover, increased life expectancy brings an emphasis on improved quality of life in old age, and other neuroscience goals include a better understanding of how to reduce and treat age-related mental impairments such as Alzheimer’s and Parkinson’s. MPE is playing a key role in much of this research, because it enables deep (1 to 1.5 mm) in vivo imaging with full 3-D resolution and low photodamage. The classic mammalian setup involves a mouse with a thin glass cover replacing a small part of the skull to provide visual access to the cortex at layers 1 through 6.
The overarching trends in neuroscience rely on faster and deeper in vivo imaging. At the same time, novel applications are being enabled by new fluorescent tools and methods, many of which use gene expression in laboratory animals ranging from insects to mice. A standout development is the new field of optogenetics, pioneered by Dr. Karl Deisseroth at Stanford University and Dr. Ed Boyden at MIT.
Optogenetics has enabled an important transition to using light not only to image structures, but also to stimulate specific cells, thanks to a class of proteins – opsins – that can be expressed in model (transgenic) animals. These proteins reside in the cell membrane and act as single-element ion gates that can be controlled by pulses of light. When irradiated at the appropriate wavelength, they enable Ca2+ or other ions to flow across the membrane, leading to a spike in the membrane potential that mimics the normal action potential used for neuron conduction and dendritic signaling processes in neural cell networks. Optogenetics has enabled researchers to progress beyond studying the structure of neural networks to provoking (or inhibiting) their activity in real time. What about monitoring the actual signals as they propagate in the neural network, typically in terms of Ca2+ spatiotemporal concentration? This also can be done using another family of genetically expressed probes that fluoresce strongly only in the presence of this ion – the so-called GECI (genetically encoded calcium indicators).
Early optogenetics research used LED light sources for wide-field photoactivation –somewhat of a “bazooka” approach. Thanks to the exquisite selectivity and penetration of MPE, an appropriate ultrafast laser can be used for highly specific optogenetics activation of particular sites, possibly in conjunction with a spatial light modulator. As shown in Figure 4, a second ultrafast wavelength can be deployed for imaging the Ca2+ activity using a GECI that is excitable at a convenient wavelength.
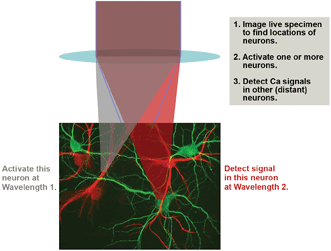
Figure 4. An advanced optogenetics experimental scenario involves MPE imaging to map the neurons, followed by activation of one or more neurons using MPE at Wavelength 1 and probing of Ca2+ activity in remote neurons by multiphoton excitation at Wavelength 2. Wavelength 1 (or sometimes Wavelength 2) is also used for the initial MPE mapping.
Higher laser power is also needed for faster imaging speed. Specifically, at present a typical multiphoton experiment with individual neuron resolution might involve stimulating and then imaging the activity of only a few neurons simultaneously. Ultimately researchers would like to simultaneously survey as many as 10,000 neurons – i.e., a column of cortex measuring 250 × 250 µm with a depth of up to 1 mm. This will require very fast scanning with fast modulators, or splitting the beam intensity and scanning the tissue with techniques such as resonant scanning, spatiotemporal multiplexing and random-access multiphoton microscopy (the latter pioneered by Dr. Peter Saggau and co-workers at Rice University). Either way, laser power will be key to obtaining acceptable signal-to-noise images at these faster sampling speeds.
Optogenetics is not the only area of neuroscience where multiple laser wavelengths are needed simultaneously. One interesting technique for mapping different neuron types is “brainbow” imaging. Here, subject animals are transfected with genes for three or more different fluorescent proteins, whose emission peaks at red, green and blue wavelengths. The various cell types in the brain express these proteins in unique stochastic ratios, each characterized by a different ratio of the fluorophores and, therefore, a unique emission spectrum and color signal. Brainbow imaging needs two or more independently tunable ultrafast sources.
Trends in ultrafast lasers driven by neuroscience needs
To better support these new imaging methods in neuroscience, laser manufacturers are focused on providing turnkey sources with higher power at longer wavelengths. In this context, the most recent advances in commercial ultrafast laser sources are in improved optical parametric oscillator (OPO) performance and a new type of ytterbium fiber laser.
A few years ago, OPOs based on fan-poled crystal technology provided a revolutionary way to generate two independently tunable wavelengths – one from the Ti:S laser and one from an OPO pumped by this laser, all under push-button integrated computer control. Dr. Emmanuel Beaurepaire of CNRS et al have recently shown that independent tunability of two synchronous pulses can excite all the brainbow expressions by using two photons from the Ti:S laser, two from the OPO and one photon each from the OPO and the Ti:S (Figure 5).
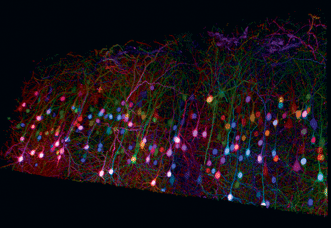
Figure 5. Trichromatic two-photon imaging of brainbow mouse cortical tissue using wavelength mixing. Courtesy of E. Beaurepaire, P. Mahou, K. Loulier, J. Livet, École Polytechnique, Palaiseau/Vision Institute, Paris. See Mahou et al (2012), Nat Methods.
The ability of the OPO to extend the long wavelength range of a Ti:S oscillator is also critical to advancing neuroscience by providing deeper images. Specifically, deeper imaging is enabled by using longer wavelengths, because this greatly reduces light scatter that usually is the depth-limiting factor. And as neuroscientists move even deeper (>1 mm) into the cortex, MPE becomes ever more challenging. A new OPO optimized specifically for very deep imaging, the Chameleon MPX OPO from Coherent, doubles the brightness of these deep images compared with earlier OPOs.
It accomplishes this by providing a combination of shorter pulses and higher average power in the optimal wavelength region (1010 to 1340 nm) for two-photon excitation of GFP and red-shifted derivatives. Moreover, this new OPO ensures shorter pulse width at the sample by means of a built-in group velocity dispersion precompensator. This is factory set to offset the typical total group velocity dispersion caused by the beam delivery, scanning and focusing optics in common commercial multiphoton microscope models.
In optogenetics, it is vital to minimize crosstalk between the stimulation and probing phases of the experiment. This is achieved by using two laser wavelengths within the same microscope setup (Figure 6). The latest Ti:S laser and OPO systems can produce the requisite paired wavelength combinations – for example, to excite a GECI (e.g., GCaMP) indicator and red-shifted opsins. The available power simultaneously available is 750 to 1000 mW at each of the two wavelengths. While this output power is perfectly adequate for conventional raster-scan MPE imaging, some advanced optogenetics and optical physiology experiments require fast point-to-point imaging techniques or simultaneous multipoint approaches that are both power hungry.
The power simultaneously available from the Ti:S laser and the OPO is about 1 W each. This output power is perfectly adequate for imaging and activating small groups of neurons. The grander goal of studying hundreds or thousands of neurons will likely require different laser types with more power or energy.
In response to these emerging “power hungry” developments in optogenetics, an ytterbium fiber laser new to the market delivers over 2 W at 1055 nm. This fiber laser enables both a decrease in overall capital equipment cost and an increase in the available power at a wavelength critical for optogenetics or signal-monitoring applications. Together with a tunable laser, this source is ideal for optogenetics with a wavelength split, such as Example 2 in Figure 6. The latest ytterbium fiber laser is one example epitomizing one company’s “industrial revolution in ultrafast lasers,” where design and manufacturing practices from industrial lasers are being deployed into state-of-the-art scientific lasers to deliver unprecedented reliability, 24/7 performance and higher overall data productivity.
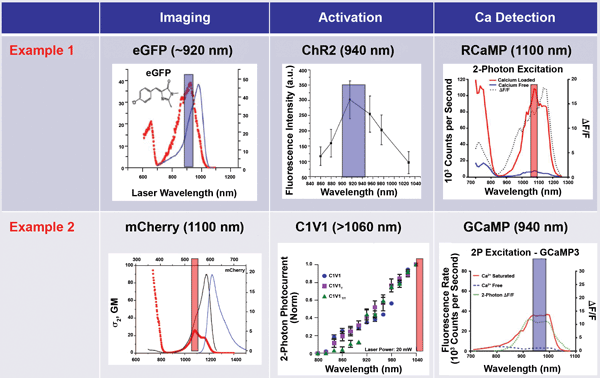
Figure 6. In optogenetics imaging, it is vital to avoid crosstalk between the activation (control) phase and the probe phase. This is achieved by wavelength separation. In Example 1, a short wavelength is used for the activation (and initial passive imaging), and a long wavelength is used for two-photon excitation of the probe. In Example 2, a long wavelength is used for imaging and activation, and a short wavelength probe is used to monitor neuron activity. Figure adapted from various sources.
While these advances are valuable advantages in the research lab, they will be absolutely vital in future applications in the preclinical and clinical markets for MPE techniques. At this time, second-harmonic generation imaging appears to be a key technique for successful translation along this path, because it avoids the use of fluorophores and genetically expressed probes that, of course, cannot be used in humans.
Summary
Laser-based microscopy continues to be one of the most valuable tools in the life sciences. As researchers develop and refine new techniques and methods, laser sources must be adapted and reoptimized to fully exploit the potential of these techniques. By closely collaborating with key researchers and institutions, laser manufacturers are responding in a timely manner with new products designed specifically for the changing needs of these applications.
Acknowledgments
The authors wish to acknowledge the assistance of Dr. Christian Eggeling (principal investigator, Human Immunology Unit, and scientific director, Wolfson Imaging Centre, Weatherall Institute of Molecular Medicine, University of Oxford, England) in the form of personal communications about optical nanoscopy techniques and specifically STED technology, including supplying Figure 1. They also wish to acknowledge the assistance of Dr. Zhen-li Huang at Wuhan National Laboratory for Optoelectronics, Huazhong University of Science and Technology, for important insights into STORM imaging and for providing Figure 3.