The mobile design featuring built-in laser scanning provides key insights into animal tissue samples and offers a window into human systems to help guide treatment.
MICHAEL E. HOLMES AND NKECHI HOLMES, TWO-LEVEL SYSTEMS, AND STEFANIE KIDERLEN AND LUKAS KRAINER, PROSPECTIVE INSTRUMENTS
For decades, optical microscopy has provided the mechanism with which to image cells and tissue for the purposes of cancer research, digital pathology, and the study of the brain. Samples are typically studied in frozen sections. Most of these systems use confocal microscopy and single-photon excitation, typically from a continuous-wave light source, to probe the sample. Multiphoton microscopy is used when deep-tissue and cellular-level resolution are desired to preserve the image of the tissue in its native environment. In laser scanning multiphoton microscopes, light from an ultrafast laser is tightly focused and scanned across the sample using fast mirrors. An image is created by detecting the fluorescence signal intensity at each point and mapping spatially with the aid of computer software.
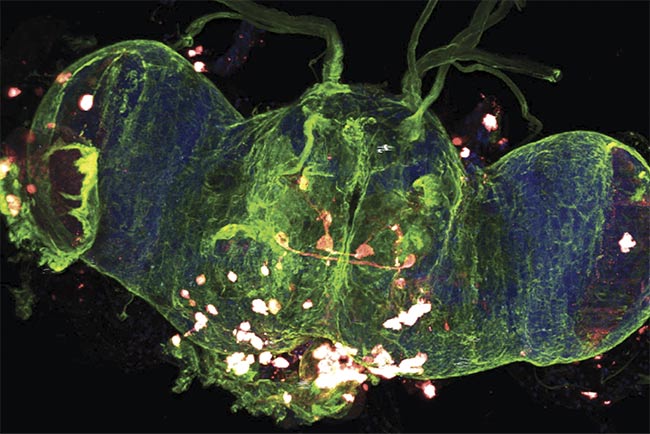
A multiphoton 3D z-stack image of a whole Drosophila (fruit fly) brain. Courtesy of Prospective Instruments.
The excitation source and signal detection implementation are critical for biological imaging, ease of use, and the uptime of the instrument. Multiphoton microscopy uses a highly focused laser beam in the near-infrared (NIR) range, typically between 780 and 1300 nm, to produce nonlinear optical effects based on the interaction of multiple photons arriving simultaneously at a molecule. Therefore, the intensity of the generated signal does not increase linearly with the number of irradiated photons but rather by the square power (for two-photon effects) or the third power (for three-photon effects). The phenomenon is confined to a very tight volume concentrated in the plane of focus, significantly reducing the absorption cross section from surrounding material. Moreover, using wavelengths in the NIR range leads to lower scattering in tissue and yields higher penetration depth compared to conventional linear confocal techniques. This allows 3D sectioning with less of the photobleaching and phototoxicity that plague single-photon techniques.
Real-time and extended-period measurements are possible — for example, in experiments studying live animals and drug target engagement. One prominent example of using laser scanning multiphoton microscopy is imaging deep in the living brain of animals (Figure 1d) by using standard markers such as GCaMP. GCaMP is a fluorophore-tagged and genetically modified calcium indicator that responds to the efflux of calcium ions (Ca2+), indicating neuronal activity. Higher penetration depth combined with lower photodamage are major advantages to using laser scanning multiphoton microscopy for living or fixed native 3D samples such as brain, embryos, live organs, or animals, or for in vitro applications such as spheroids or organoids.
Finding versatile solutions
As the applications for multiphoton microscopy grow, so does the demand for an easy-to-use, transportable, and multimodal tool to adapt to any indoor working space. An ideal system should produce high-quality results and process high-volume data quickly while providing a powerful time-saving workflow. Also, the capability to perform multimodal analysis on the same region of interest without the inconvenience of moving the sample to another system maximizes content and provides more powerful complementary insights.
Rugged and industry-ready devices that can be used for clinical research and are capable of operating in any indoor environment are likely the future of laser scanning multiphoton microscopy. Commercially available systems lack the agility and mobility to be used outside of the laboratory. The scan head (including beam-steering optics) and ultrafast laser source design are critical to the overall functionality and usability of the microscope. Most systems separate the two components, and they are typically manufactured by separate suppliers.
The industry standard for the femtosecond laser system has been titanium:sapphire (Ti:sapphire) laser technology for nearly 40 years. These systems are very complicated to operate, typically requiring a trained expert, and they pose a significant safety risk. They require a large optical table, a chilled water supply, and special infrastructure, and they are fragile. Their size and weight and the amount of power they consume make them inappropriate for most clinics, and they are impossible to transport. Clinical researchers need a portable and turnkey multimodal microscope that can be incorporated in any indoor environment to be usable by most staff.
Next-gen instrumentation
The design of an ultraflexible laser scanning multiphoton microscope for in vivo imaging is predicated on a highly flexible front end and laser design. The ability to easily maneuver the scan head is key to allowing adaptability to a broad variety of setups, from slides to even live mice. The design of the scan head is crucial to enabling the microscope to be adaptable to almost any experiment. It should include the galvo-resonant scanners, photomultiplier tubes, objectives, wide-field camera and illumination, complete scan-path optics, and lasers — all in one compact, ready-to-go, modular form factor (Figure 1)1.

Figure 1. An ultrafast air-cooled fiber laser engine is integrated into the free-moving scan head of the MPX-1040 microscope (a). The microscope consists of two parts (b, c) that are connected through a flexible umbilical cord. Example uses include live-mouse multiphoton imaging (d), whole organ multiphoton imaging of a mouse kidney (e, left) and colon (e, right), and imaging of standard 2D samples mounted on an objective slide (f). Courtesy of Prospective Instruments.
The power and size of the laser source are critical in the design of a truly portable and agile laser scanning multiphoton microscope. Fiber lasers are well suited for this purpose because they are compact, rugged, and energy-efficient for almost any indoor workspace. They are air-cooled and maintenance-free; therefore, there is no need for a chilled water supply or service. For example, the MPX-series microscope, recently developed by Prospective Instruments, is engineered with a dual-wavelength femtosecond fiber laser with a fixed output at 1040 nm (>500 mW) and a second widely tunable laser in the range of 750 to 960 nm and 1150 to 1300 nm (>200 mW). The pulse duration is <140 fs when operating at 80 MHz repetition rate. These parameters are all measured at the sample position and are sufficient for sectioning >1 mm.
High-resolution multiphoton imaging has been demonstrated on animals, including zebrafish and the fruit fly (Drosophila). Both provide invaluable information for understanding and advancing human pathophysiology. In recent experiments, both excitation (130 fs at 1040 nm) and detection signals were imaged through a 20× water objective (NA 1.0).
The fixed Drosophila samples were immunohistochemistry (IHC) stained with fluorophores (Hoechst and Alexa-Fluor 488), whereas the living zebrafish larvae expressed fluorophore-tagged proteins (green fluorescent protein, or GFP, and mCherry).
Structural and functional details were observed using complementary two-photon and second-harmonic generation (SHG) imaging (Figure 2) with z-stack and overlayed images of a living zebra-fish showing differentiated features such as the beating heart. Another example, in Figure 3, shows features of the muscle (z-bands) and a migrating cell, with dual modality overlayed. SHG is a label-free method of differentiating collagen and muscles from the surrounding environment. Contrast from colors enhances the image specificity that is necessary for understanding the relationship between healthy, damaged, and diseased tissue, or between different cell types and extracellular matrix components.
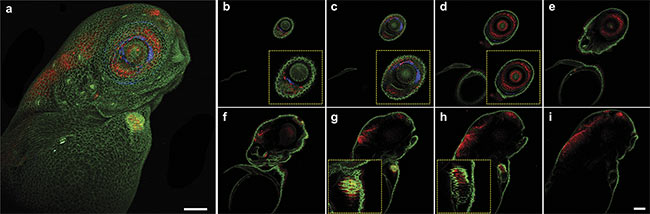
Figure 2. Multiphoton 3D z-stack imaging of a living zebrafish larva, expressing alpha-catenin-YFP (yellow fluorescent protein) (green) and tp1:mCherry-NLS (nuclear localization signals) (red). Maximum intensity projection of a 510-µm z-stack with a step size of 10 µm — second-harmonic generation (SHG) (blue) and two-photon channels (green and red) (a). Z-layers going from 20 µm (b) to 300 µm (i) in 40-µm steps. Close-ups depicting specific features such as the eye (b-d) or the beating heart of the zebrafish larva (g, h). Scale bar: 100 µm. Courtesy of Prospective Instruments.
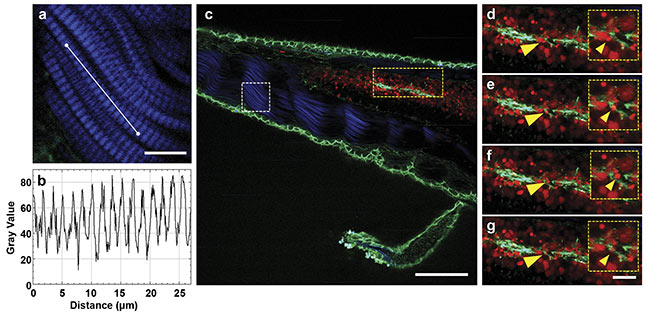
Figure 3. Multiphoton time-lapse imaging of a living zebrafish larva, expressing alpha-catenin:YFP (green) and tp1:mCherry-NLS (red). A close-up of the SHG signal (blue) originating from the muscles (a) and the respective intensity plot to distinguish the z-bands (b). An overlay of the SHG signal (blue) and the two-photon channels for YFP (green) and mCherry (red) expressed in the cell-cell contacts and the nuclei of cells in Notch-expressing tissues, respectively (c).
Close-ups of different timepoints, with 1-min steps in between. Migrating cells (yellow arrows, d-g). Scale bars: 10 µm (a), 100 µm (c), and 20 µm . Courtesy of Prospective Instruments.
Zebrafish imaging
The zebrafish has long served as an invaluable subject in the fields of metabolic pathology, developmental biology, and neuroscience. Its status as a vertebrate, as well as its ability to lay large numbers of collectible, transparent eggs, and its sustained transparency in the larval stage make it a highly practical and expedient live model for research. The genetic composition of zebrafish shows about 70% homology with human genes and a high sensitivity to pharmacological and genetic manipulations2, allowing it to serve as a model for understanding various neurological disorders such as Parkinson’s disease, Alzheimer’s disease, and epilepsy, as well as psychiatric disorders, including attention deficit hyperactivity disorder and schizophrenia.
Imaging the zebrafish brain does not capture the full psychosocial complexity of the human brain, as the fish’s neurological circuit system contains fewer neurons, and there is linear organization of brain regions. But its optimal translucency allows its full volume to be captured in one image with maximized clarity. As displayed in Figure 3, the use of multiphoton imaging enhances these features, as the reduced phototoxicity allows live imaging of the zebrafish from its neural network down to its subsynaptic components3.
This is especially beneficial for retinal imaging. The zebrafish has a vertebrate-like retina with a complex and self-contained network of neuron populations. Overall, there are five different types of neurons in the retina: photoreceptors, bipolar cells, retinal ganglion cells, amacrine cells, and horizontal cells. The ganglion cells are the only ones that fire action potentials, while photoreceptors are the only neurons that respond directly to light. There are also two types of photoreceptors, called rods and cones. Rods are large and cylindrical and much more sensitive to light. Cones are shorter and are solely responsible for color vision. The cones have subtypes that contain photopigments that are sensitive to different wavelengths. Incorporating genetic techniques with the help of advanced multiphoton microscopes can enhance our understanding of retinal development, morphology, and diseases such as macular degeneration4.
In the referenced zebrafish studies, the living larvae were treated with tricaine to immobilize the sample. Then they were mounted in 0.8% agarose in a 35-mm petri dish. When the agarose was polymerized, the larvae were covered with 0.5× E2 medium and imaged. Near-field 3D imaging (Figure 4) allows detailed visualization of neurons in the eye within the anterior region of the body. Utilizing the far-field magnification setting allows visualization of neurons running along the lateral line. It is also possible to use the near-field setting to create a high-resolution image of the retina and its neurons. Living zebrafish specimens, expressing fluorophore-tagged proteins, yield an intense and robust fluorescence signal, which therefore helps mitigate photobleaching. Time-lapsed migrating cells were imaged with filopodia over an extended time. Additionally, these changes in magnification permit stacking and animation of multiple images, providing additional visualization of the fish’s musculature and vasculature that would not be produced using a traditional confocal microscope.
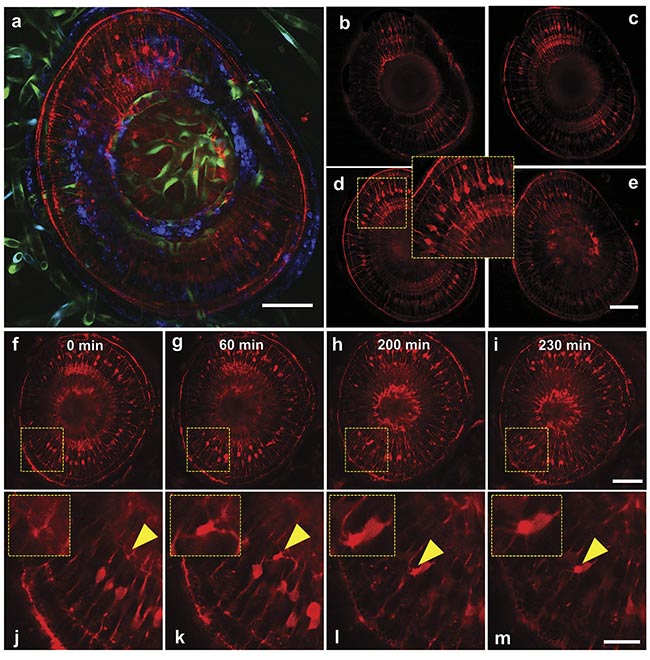
Figure 4. Multiphoton 3D z-stack imaging (a-e) and time-lapse imaging (f-m) of an eye of a living zebrafish larva, expressing tp1:LifeAct-mCherry (red) and Kdrl:GFP (green fluorescent protein) (green). Maximum-intensity projection of a 200-µm z-stack with a step size of 20 µm — SHG (blue) and
two-photon channels (green, red) (a). Z-layers going from 20 µm (b) to 80 µm (e) in 20-µm steps. Timelapse of a zebrafish eye (f-i) with the respective close-ups showing a migrating retina cell (j-m). Scale bars: 50 µm (a-i) and 20 µm (j-m). Courtesy of Prospective Instruments.
Imaging Drosophila brain
Drosophila melanogaster is another important organism whose neuroanatomy has been studied using multiphoton microscopy. The focused imaging of the common fruit fly has revealed several insights into the function of the olfactory system, neuroactivity, and neuronal tracing. This area of neuroscience focuses on the influence that different neuron populations have on each other and the functional connection between the anatomical regions of the brain. Additionally, the fruit fly has been used as a model in the study of neurodegenerative diseases, given that over half of the genes associated with neurodegeneration in Drosophila brain have counterparts in mouse or human brains5. The fruit fly also possesses several practical advantages, including its small size, rapid generation cycle, and ability to produce large numbers of genetically modifiable larvae6.
The olfactory system of Drosophila melanogaster allows it to recognize hundreds of distinguished odorants. Olfaction plays a critical role in allowing these insects to find shelter and food, and to mate. Olfactory receptor neurons detect and transmit chemical signals to the central nervous system. The process occurs within the fruit fly’s primary olfactory sensory organs, the antennae, and the maxillary palps. On the surface of these organs reside sensory hairs called sensilla, which are connected to the dendrites of the olfactory receptor neurons and covered in sensillar lymph, a clear fluid containing several types of odorant-binding proteins7.
The purpose of examining this network of neuron populations is to investigate the relationship between odor perception and behavior. In one study conducted by a research group at Stanford University, the antennae of the fruit fly were stained to visualize its neurons and compare the development of the olfactory neural circuit ex vivo with the in vivo conditions of the sample using time-lapsed images of the cells. One of the conclusions reached in the study was that the olfactory circuit in the explant culture develops similarly to the way it does in vivo but at a slower rate8. The use of two-photon microscopy has allowed researchers to differentiate between different populations of neurons in the entire brain and to simultaneously monitor their activity in 3D.
The whole Drosophila brain was resected, and the cell nuclei were stained with DAPI (4′,6-diamidino-2-phenylindole). The fruit fly was genetically modified to express different fluorophores, allowing easier differentiation between the neuron populations. An overlay of these stained cells is shown in Figure 5, while Figure 6 serves as its orthogonal counterpart, allowing complete 3D visualization of the brain along all planes. The nervous system is a complex network of various cells working in conjunction with each other. Imaging the different neuron cells all at once in their 3D environment can allow the ability to see how they interact with each other or their environment.
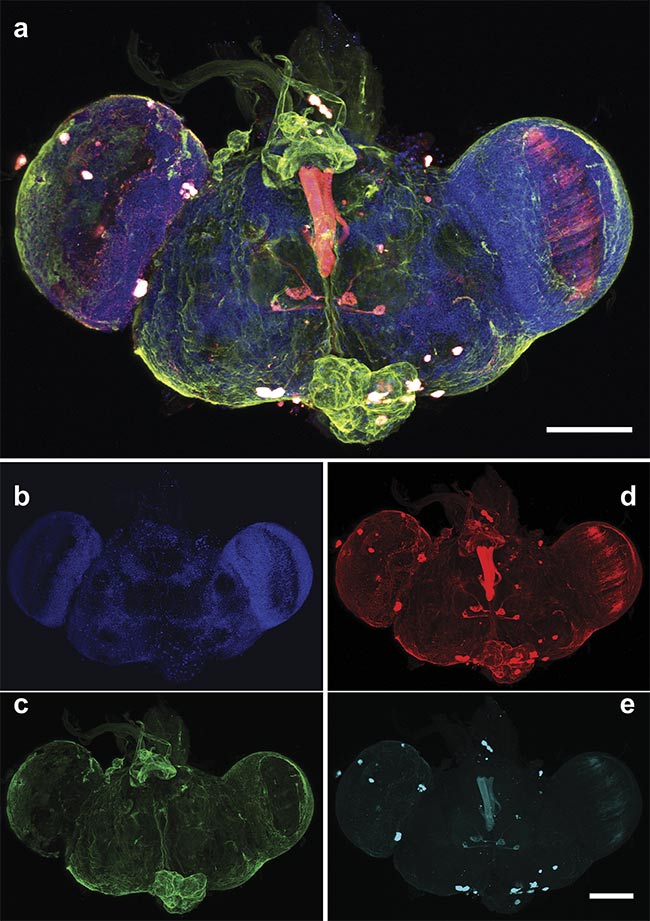
Figure 5. A multiphoton 3D z-stack of a whole Drosophila brain. An overlay of the cell nuclei (Hoechst) (blue), different neuron populations — AF568 (red) and AF647 (cyan) — and the SHG signal (green). An overlay of the maximum intensity projection of a 190-µm z-stack with a step size of 2 µm (a) and the respective single two-photon (b, d, e) and SHG (c) channels. An Olympus 20× water objective (NA 1.0) was used. Scale bars: 100 µm. Courtesy of Prospective Instruments.
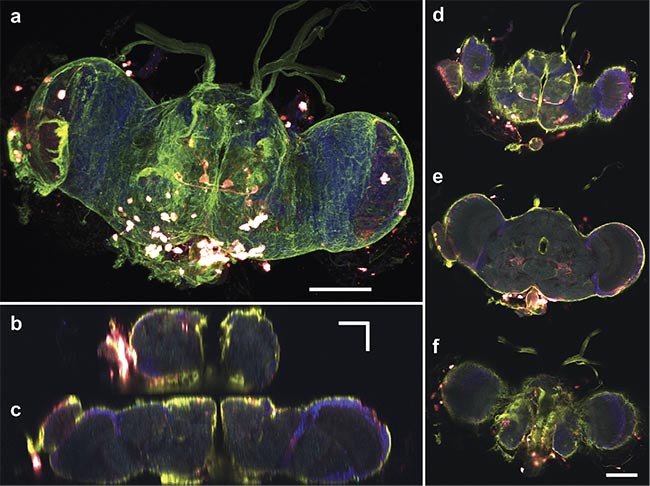
Figure 6. A multiphoton 3D z-stack of a whole Drosophila brain. An overlay of the cell nuclei (Hoechst) (blue), different neuron populations — AF568 (red) and AF647 (cyan) — and the SHG signal (green). A maximum-intensity projection of a 132-µm z-stack with a step size of 1 µm (a) and the respective orthogonal views in y,z (b) and x,z (c). Top, middle, and bottom layers (d-f), respectively. An Olympus 20× water objective (NA 1.0) was used. Scale bars: 100 µm (a, d-f) and 50 µm (b, c). Courtesy of Prospective Instruments.
Neuroscientists see the importance of studying the entire brain and not just individual regions. It could be shown, however, that even in the resting state many brain areas are active simultaneously in Drosophila. For example, Maxwell H. Turner and his colleagues have shown that several neurons in one region can have an impact on another region9. Overall, the fruit fly maintains good conservation of neural signaling pathways and overall cellular processes, which means there is a lot to be learned through studying this organism. This conservation has allowed biomedical researchers to not only image the fruit fly’s anatomy, but also to assess the effects of certain treatments and drugs without the risks or ethical ramifications of testing on humans. For this type of assessment, it is essential to be able to image the whole brain in its full depth.
The use of laser scanning multiphoton microscopy is quickly growing throughout clinical and biological research for its superior benefits to traditional optical imaging. The instruments have helped deepen the understanding of various diseases, neuronal functionality, and structural relationships, as well as developmental processes. Until now, limitations on traditional instrumentation have slowed progress in mainstream research and industrial applications. However, the next generation of multiphoton microscopy systems will have the power to revolutionize biomedical research and overcome conventional barriers that are limiting access to deep-tissue imaging. Future developments and improvements to the platform include additional modalities such as three-photon and fluorescence lifetime imaging, improved optical performance using adaptive techniques, and a faster and more intelligent imaging workflow. And the systems could very well accomplish all of these improvements while maintaining the same physical form, efficiency, and agility.
Meet the authors
Michael E. Holmes, Ph.D., is founder of two-level systems, a distributor of novel scientific laser and microscopy solutions and the exclusive U.S. partner of Prospective Instruments. He received his doctorate in physics, with a research focus on ultracold atoms and molecules, from the University of Rochester. He has held various executive roles within the scientific laser and cryogenic industries; email: [email protected].
Nkechi Holmes is an undergraduate student majoring in biochemistry and neuroscience at the University of Pennsylvania. She also serves as a marketing research assistant at two-level systems; email: [email protected].
Stefanie Kiderlen, Ph.D., is senior applications scientist and expert in-house microscopist at Prospective Instruments. She received her doctorate in biophysics, with a research focus on cell biological imaging, from Ludwig-Maximilians-University of Munich; email: [email protected].
Lukas Krainer, Ph.D., founder and CEO of Prospective Instruments, provides the creative inspiration and intellect for this new philosophy of turnkey and portable multiphoton imaging; email: [email protected].
Acknowledgments
The authors are grateful to researchers Sara Caviglia, Ph.D., and professor Stephan Neuhauss, Ph.D., from the University of Zürich, for their generous contributions of zebrafish samples.
References
1. M.E. Holmes et al. (2022). Next-generation laser scanning multiphoton microscopes are turnkey, portable, and industry-ready. Microsc Today, Vol. 30, No. 3, pp. 16-23,
www.doi.org/10.1017/s1551929522000657.
2. K. Howe et al. (2013). The zebrafish reference genome sequence and its relationship to the human genome. Nature, Vol. 496,
No. 7446, pp. 498-503, www.doi.org/10.1038/nature12111.
3. L.C. Leung et al. (2013). Imaging zebrafish neural circuitry from whole brain to synapse. Front Neural Circuits, Vol. 7,
www.doi.org/10.3389/fncir.2013.00076.
4. S. Niklaus and S.C.F. Neuhauss (2017). Genetic approaches to retinal research in zebrafish. J Neurogenetics, Vol. 31, No. 3, pp. 70-87, www.doi.org/10.1080/01677063.2017.1343316.
5. D. Lessing and N.M. Bonini (2009). Maintaining the brain: insight into human neurodegeneration from Drosophila melanogaster mutants. Nat Rev Genet, Vol. 10, No. 6,
pp. 359-370, www.doi.org/10.1038/nrg2563.
6. N. Tolwinski (2017). Introduction: Drosophila — a model system for developmental biology. J Dev Biol, Vol. 5, No. 3, p. 9, www.doi.org/10.3390%2fjdb5030009.
7. L.B. Vosshall (2000). Olfaction in Drosophila. Current Opin Neurobiol, Vol. 10, No. 4, pp. 498-503, www.doi.org/10.1016/S0959-4388(00)00111-2.
8. T. Li et al. (2021). Cellular bases of olfactory circuit assembly revealed by systematic time-lapse imaging. Cell, Vol. 184,
No. 20, pp. 5107-5121, www.doi.org/ 10.1016/j.cell.2021.08.030.
9. M.H. Turner et al. (2021). The connectome predicts resting-state functional connectivity across the Drosophila brain. Curr Biol, Vol. 31, No. 11, pp. 2386-2394, www.doi.org/10.1016/j.cub.2021.03.004.