By Marco Arrigoni, Darryl McCoy and Nigel Gallaher, Coherent, Inc.
After years of domination by a single ultrafast laser technology (Ti:S) and
its associated add-ons,
laser manufacturers have
broadened their technology portfolios to better support a diversified and dynamic
nonlinear imaging
applications space.
Multiphoton excitation (MPE) microscopy is one of the most powerful imaging tools for life science research and preclinical applications because it provides inherent 3-D resolution with minimal damage to live samples. In multiphoton microscopy, an ultrafast laser is tightly focused at a target depth in the sample and scanned in the X-Y plane defined by this depth. Only in this plane is peak intensity high enough to drive two-photon or other nonlinear interactions resulting in a signal sufficient to construct an image. The continued diversification of applications for multi-photon microscopy, particularly in the area of neuroscience, has created a need for a broader portfolio of turnkey laser sources. To that end, the latest ytterbium-based laser systems complement existing MPE laser sources based on time-proven titanium:sapphire (Ti:S) technology and its accessories.
Neuroscience and optogenetics
The field of neuroscience has become the key application area for multiphoton optical microscopes because of its growing worldwide funding, both institutional and private. Plus, multiphoton microscopy offers the unique ability to produce high-resolution neuronal images in the cortex of living brains. In particular, the technique of optogenetics is enabling new understanding at the level of individual neurons, correlating them with function in the neural networks of model animals.
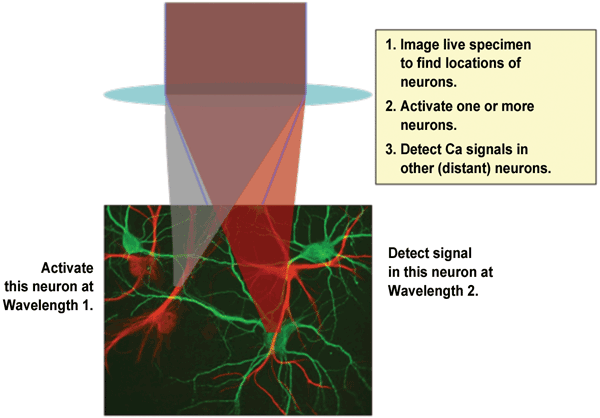
Figure 1. In a typical optogenetics experiment, two different wavelengths are required: one for
photoactivation of target neuron(s) and a second wavelength to detect Ca2+
ion spikes in interconnected neurons. The detection (probe) laser also
is used at the start of the experiment to map the structure of the
target tissue before activation. Courtesy of Coherent Inc.
Pioneered by Dr. Karl Deisseroth’s group at Stanford University and Dr. Ed Boyden’s group at MIT, optogenetics is an innovative method where light is used to stimulate the opsin proteins of specific cells. When irradiated at the appropriate wavelength, these membrane-resident proteins enable Ca2+ (or other) ions to flow across the membrane, leading to a spike in membrane potential that mimics the normal action potential used for neural signaling. Optogenetics uses light, often (but not always) in conjunction with microscopy, to activate or silence neural networks in the brain cortex of live animals that have been genetically modified to express these light-sensitive probes.
An optogenetics experiment on the
microscopic scale typically involves first imaging the neurons, then measuring signaling activity (including timing) in the local neural network, following the stimulation or silencing of other parts of the network. Signaling is usually measured in terms of local Ca2+ ion concentration using another type of genetically encoded probe, such as GCaMP, that fluoresces more strongly in the presence of Ca2+ ions when activated with a laser beam. The combination of these protocols is referred to as all-optical physiology. Ultrafast lasers for MPE are ideal for all three protocols (imaging, stimulation/silencing and detection) because MPE enables deep imaging (1-1.5 mm) with full 3-D resolution and low photodamage. These advantages result from the minimized scatter and negligible one-photon absorption at the long (NIR) MPE excitation wavelengths. As a result, larger brain volumes can be studied in vivo for extended periods.

Figure 2. Higher power from femtosecond laser sources
enables simultaneous activation of multiple neurons within a population.
Rendering courtesy of the Hausser Lab, University College London.
The classic experimental setup involves a mouse with a thin glass cover replacing a small part of the skull to provide visual access to the cortex at layers 1 through 6. To obtain useful data, it is important to minimize crosstalk between the stimulation and signal-detecting phases of the experiment. This typically is achieved by using two laser wavelengths within the same microscope setup. Figure 1 schematically illustrates the key principles of a typical optogenetics experiment. Cell morphology may be mapped first by either the probe or the activation wavelength, followed by activation at selected locations using an appropriate laser wavelength in conjunction with random-access scanning or patterned illumination. Signaling activity – as indicated by transient local Ca2+ ion concentration – is probed simultaneously by scanning the sample, usually with a second wavelength.
Dual-wavelength output
Owing to the growing range of calcium indicators and optogenetics probes, the
choice of optimal wavelengths is somewhat fluid and depends on a number of
factors. The current trend is toward longer wavelengths, which minimize optical damage and maximize imaging and activation depths. Longer-wavelength calcium indicators, like RCaMP, are progressing in sensitivity but are not yet widely adopted or available. Fortunately, some longer-wavelength optogenetics activators already are well developed and characterized for multiphoton use. An example is C1V1, which can be effectively activated in the 1040- to 1080-nm range. This wavelength also can map neural structures using a fluorescent protein such as one of the mFruits. Probing could then be done at 920 nm with the fluorescent calcium indicator GCaMP3, which has low sensitivity in the 1040- to 1100-nm region.
It is common practice to run calcium activity and photoactivation processes simultaneously so as not to lose precious
data from neural kinetics. Care and empirical analysis of crosstalk between wavelengths are therefore paramount to ensure that no unwanted action potentials are generated – for example, by Ca2+ image scanning methodologies. Given that two separate wavelengths are routinely required, a few laser system combinations are possible. Two Ti:S lasers can attain the correct wavelength pairings, as can a Ti:S laser pumping an optical parametric oscillator (OPO). The latter solution provides two independently tunable wavelengths that surpass a Ti:S laser’s tuning range.
Even so, higher power often is desirable both for enabling high-speed calcium imaging with low signal-to-noise ratios and for modern photoactivation techniques. Today’s experiments might simultaneously excite a few tens of neurons with scanless imaging methodologies, requiring lasers with high peak power in both beams.
A first alternative solution to the quest for power came with the introduction of an ytterbium fiber laser (the Fidelity from Coherent), a simple industrial-grade laser offering >2 W of fixed-wavelength output near 1050 nm and a short pulse width (<70 fs). When coupled with a Ti:S laser at 940 nm, this combination greatly increased the number of neurons that could be stimulated, as noted by researchers in our October 2014 BioPhotonics article “Optogenetics Advances Ultrafast Lasers.”
A more streamlined solution for many optogenetics applications is the latest generation of dual-wavelength ytterbium-based lasers incorporating a completely integrated and software-tunable OPO in a single compact box. An example is the Chameleon Discovery from Coherent. The primary output of this laser is continuously tunable from 680 to 1300 nm,
with short pulse widths (100 fs) and power up to 1.4 W, while simultaneously providing 1.5 W of output at 1040 nm. Moreover, because shorter pulse widths are believed to render brighter images, this new laser includes an internal and fully automated group velocity dispersion precompensator, enabling the shortest possible pulse width at the sample. This high-power combination is suitable for the typical long-wavelength optogenetics scheme previously described. These two embodiments of ytterbium technology already have previously been reported to increase the number of activated neurons. Looking to the immediate future in optogenetics, these new widely tunable lasers are also well suited to optogenetics research based on alternative wavelength pairings, such as those employing red-shifted calcium indicators of the RCaMP family currently under development.
The wide tuning range, high power and short pulses of the primary output are also suitable for optimum excitation of all popular fluorophores (inclusive of Alexa Fluor dyes and calcium indicators, as well as conventional and red-shifted fluorescent proteins). While optogenetics may represent the newest and hottest area of multiphoton applications, a number of other multiphoton experiments may also benefit from this new type of dual-wavelength laser. These include multimodal studies, where two types of imaging methods are used simultaneously or near-simultaneously. Examples include using the same 940- and 1040-nm wavelength pairing for dual-protein expression to image two transgenic proteins, such as GFP and mFruits, without crosstalk.
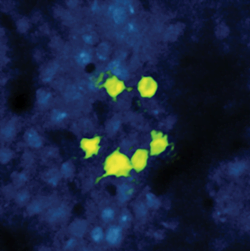
Figure 3. Neurons in the cortex of a mouse express
proteins enabling the ‘reading’ and ‘writing’ of electrical activity.
Six neurons arranged in the shape of a smiling face were simultaneously
activated with light. The response of those neurons is color-coded in
green, indicating (via expression of GCaMP6) successful activation of
this neuronal pattern of activity. Courtesy of Lloyd Russell, Hausser
Lab, UCL.
Another application is using the fixed 1040-nm output for imaging mFruits while using the tunable output in the
800- to 900-nm range to simultaneously excite “classical” dyes or to image structures using second- or third-harmonic- generation imaging. Yet another option is to use 1040 nm for two-photon excitation of mFruits, together with a longer tunable wavelength (e.g., 1250 nm) for second- or third-harmonic generation, or even three-photon excitation of some green-emitting fluorophores.
Another group of imaging techniques requires two different ultrafast wavelengths that must be phase correlated; i.e., they must originate from the same laser source. This is the case for stimulated Raman scattering or coherent anti-Stokes Raman scattering, both of which are used to probe moieties such as lipids without the need for exogenous markers. The inherent synchronization of the primary and secondary output of these new dual-wavelength ytterbium lasers make them an excellent choice for such techniques.
Ti:S lasers for deeper images
Despite undeniable advantages of these dual-wavelength ytterbium lasers, manufacturers do not yet view them as a universal solution for all multiphoton microscopy research. (And, judging by continued healthy sales of Ti:S-based lasers, neither do end users.) Rather, they are important new arrows in a diverse quiver of multiphoton microscopy lasers. To understand why market interest remains so strong, it is useful to examine the advantages of established Ti:S-based laser solutions.

Figure 4. The latest
dual-wavelength lasers, such as the Chameleon Discovery from Coherent,
are suitable for optogenetics and other microscopy applications that
require ultrafast pulses at more than one wavelength. Courtesy of
Coherent Inc.
For researchers needing only a single tunable output, Ti:S lasers are the highest-power sources of tunable femtosecond pulses for MPE applications, with up to 3.5 W of average power. Moreover, because of their maturity, some of these lasers also offer a lower capital cost than new dual-wavelength ytterbium lasers. With pulse-width options shorter than
75 fs and optional variable group velocity dispersion precompensation, Ti:S lasers deliver both the highest average power and the highest peak power at the sample. High power enables deeper imaging, which is important to all live organism/tissue imaging. Higher power also provides maximum image intensity to enable faster imaging using either multipoint scanning methods or the latest high-speed microscope scanning systems based on acousto-optic modulators.
Ti:S plus independent
OPO tuning
Another advantage of Ti:S lasers is
the option to add an integrated (or stand-alone) OPO, which extends the tuning range into the NIR and provides synchronized dual-wavelength output. Unlike the new ytterbium lasers, these two wavelengths are independently tunable, thanks to a powerful OPO technology called fan-poled crystals, which David Armstrong described in “Ultrafast Lasers for Microscopy: Flexibility Plus State-of-the-Art Performance,” in the January 2011 issue of BioPhotonics. The ability to generate two independently tunable wavelengths at a power level of ~1 W adds unparalleled flexibility to potential experiments.
Ti:S plus 1055-nm ytterbium
The range of available options for advanced neuroscience and other multiphoton imaging also includes another pairing of laser sources, as mentioned above. At their full power, Ti:S lasers can produce over 1.5 W at 940 nm. Combined with the 2-W output of a short-pulse fiber laser, researchers can have the highest signal excitation sources for dual-wavelength optogenetics and other studies. This modular approach may also be preferable in multiuser facilities where some researchers are working in optogenetics. Other users may just want high power for deeper second-harmonic-generation images.
Meet the authors
Marco Arrigoni is director of marketing at Coherent Inc.; email: [email protected]. Darryl McCoy is a director
of product marketing; email: [email protected]. Nigel Gallaher is a director of product marketing; email: [email protected].